Nouvelles occurrences de Modulidae (Mollusca: Gastropoda)
dans les gisements européens de l'Éocène, de l'Oligocène et du Miocène:
données des collections du XIXe siècle
Carnets Geol., Madrid, vol. 20, no. 7, p. 125-139.
Pierre Lozouet, Bruno Cahuzac, Laurent Charles
(traduction post-éditée par N. Bacaër, suggestions d'amélioration : nicolas.bacaer@ird.fr)
Résumé
L'examen de collections historiques déposées au Muséum d'Histoire Naturelle de Bordeaux et à la Faculté des Sciences de l'Université de Bordeaux a permis d'identifier de nouveaux taxons de Modulidae dans l'Oligocène et le Miocène du bassin aquitain. Ces Modulidae appartiennent aux groupes américains Modulus modulus et Trochomodulus. Trois nouvelles espèces sont décrites : Modulus benoisti nov. sp. (Serravallien), Trochomodulus stampinicus nov. sp. (Rupélien), proche de Trochomodulus sublaevigatus (Orbigny, 1852), une espèce chattienne, et Incisilabium trochiformis nov. sp. (Priabonien). Ce dernier est le plus ancien Modulidae connu avec Incisilabium parisiensis (Deshayes, 1832) (Éocène moyen) précédemment classé dans les Trochidae. Ces résultats montrent que la paléobiogéographie et l'origine de la famille des Modulidae doivent être réexaminées.
1. Introduction
Les Modulidae sont une famille de cérithioïdes littoraux qui se rencontre dans les régions tropicales et subtropicales. Il est relativement rare dans les gisements fossiles. C'est aussi une famille paucispécifique avec environ 12-16 espèces vivantes. Les régions tropicales américaines sont relativement riches avec au moins onze espèces vivantes, alors qu'il y a deux ou trois espèces dans le Pacifique oriental et neuf espèces ou plus dans l'Atlantique occidental, et un nombre équivalent ou plus d'espèces fossiles (Landau et al ., 2014). En comparaison, seules deux espèces sont reconnues dans la vaste région ouest-indo-pacifique (Lozouet & Krygelmans , 2016 ; Bouchet , 2018), et une seule espèce est connue des gisements du Miocène européen.
Cet article rapporte la redécouverte de Modulidae dans l'Oligocène et le Miocène du Bassin d'Aquitaine, appartenant à des genres que l'on pensait auparavant exclusivement caribéens (Modulus et Trochomodulus), dans trois collections historiques (MHNBx: coll. Benoist ; UB: collection générale et coll. Grateloup). Il décrit également une nouvelle espèce d' Incisilabium de l'Éocène supérieur.
Abréviations utilisées: Longueur, SL; Longueur de la dernière verticille, SWL; Diamètre, SD (mesures de la coque); MNHN-IM, Muséum national d'Histoire naturelle (Muséum national d'histoire naturelle), Malacologie; MNHN.F, Muséum national d'Histoire naturelle , Paléontologie; MHNBx, Muséum d'Histoire naturelle de Bordeaux (Muséum d'histoire naturelle de Bordeaux); UB, Université de Bordeaux (Université de Bordeaux), no. TYFIPAL [" TYpes et FIgurés en PALéontologie " (TYpe et spécimens FIgured en PALéontologie)].
2. Cadre géologique et collections historiques de Benoist
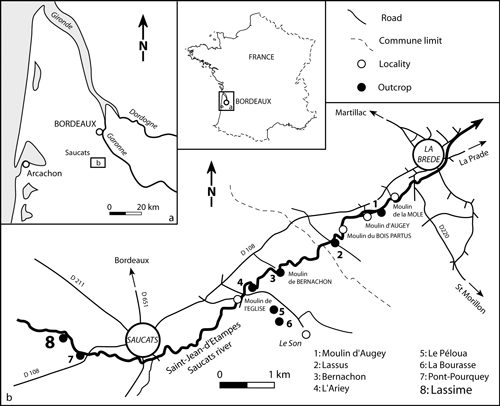
Le long du ruisseau "St-Jean-d'Étampes" (Gironde, Saucats et La Brède), des strates géologiques du Miocène inférieur au Miocène moyen sont exposées (Fig. 1 ). Il s'agit notamment des affleurements classiques du stratotype aquitain et des affleurements du stratotype burdigalien. Ce territoire est partiellement protégé et constitue depuis 1982 la " Réserve Naturelle Géologique de Saucats-La Brède " (Réserve Géologique Naturelle de Saucats-La Brède, RNG), qui fut la première Réserve Géologique créée en France. La plupart des strates exposées appartiennent au Miocène inférieur (Aquitanien et Burdigalien) mais une partie limitée de la section, sur le site nommé "La Sime, Cazenave" (maintenant appelée Lassime), contient des couches appartenant au Miocène moyen marin (Serravallien) (Mayer , ; Linder , 1872). Les dépôts serravalliens recouvrent les niveaux les plus élevés du Burdigalien local (Fig. 2 ). Cette section montre donc qu'il existe un écart stratigraphique régional correspondant à un hiatus incluant les stades du Haut-Burdigalien, du Langhien et du Serravallien inférieur (Londeix & Jan Du Chêne , 1998 ; Parize et al ., 2008).
Figure 1 : Carte de localisation de la zone Miocène de Saucats, exposée entre les villages de La Brède (en aval) et de Saucats (en amont). Succession des affleurements stratotypiques de l'Aquitanien: 1 à 4; Burdigalian: 5 à 7; Serravallien: 8 (section Lassime).
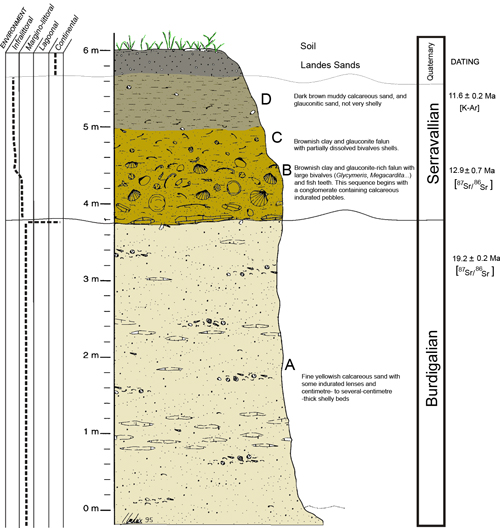
Figure 2 : La coupe de Lassime. La datation a été effectuée par les isotopes du strontium [ 87 Sr / 86 Sr] (Cahuzac et al. , 1997) dans les coquilles et la datation K-Ar des grains de glauconite non retravaillés (niveau D) (Parize et al., 2008). Modifié d'après Londeix et Jan Du Chêne (1998).
La macrofaune de ce gisement, et en particulier la malacofaune, a été répertoriée il y a longtemps par Benoist (1873 , 1874) qui a identifié 230 espèces. Benoist (1845-1904), dentiste de profession, est l'un des principaux contributeurs aux connaissances actuelles sur les mollusques fossiles de la vallée de Saucats. Sa collection a été donnée et en partie conservée au Muséum de Bordeaux depuis 1900.
L'excavation d'un nouvel affleurement sur le site de Lassime (sur la rive droite du ruisseau) a été réalisée par la RNG. Au stade serravallien, trois unités ont été distinguées. La séquence du Miocène moyen commence par un conglomérat transgressif contenant des galets calcaires. La datation utilisant les isotopes du strontium sur des coquilles de Glycymeris (Bivalvia) a fourni un âge de 12,9 ± 0,7 Ma pour cette couche basale (Fig. 2 , niveau B, Cahuzac et al. , 1997). En utilisant des grains de glauconite non retravaillés du haut de l'affleurement, la méthode K-Ar a donné un âge estimé de 11,6 ± 0,2 Ma (Fig.1 , niveau D). Malheureusement, les mollusques sont mal conservés dans cet affleurement, à l'exception des coquilles très épaisses et des gros bivalves à coquille calcitique (Pectinidae). Néanmoins, les collections nous ont permis de compléter nos connaissances sur les Pectinidés et de lister 10 espèces (Cahuzac , 2005).
Cossmann et Peyrot (1909-1924) et Peyrot (1925-1935) ont répertorié 135 espèces du Serravallien de Lassime. Parmi eux, il y a 25 spécimens typés et figurés. Cependant, Cossmann et Peyrot indiquent pour 71 espèces " fide Benoist ", ce qui signifie clairement qu'ils n'ont pas personnellement vu la plupart des espèces. Cahuzac et Cluzaud (1999a , 1999b) n'ont trouvé que 15 espèces dans les anciennes collections. Pour ces auteurs, un doute existe sur la présence de nombreuses espèces à Lassime. La question est de savoir si BenoistLes listes (1873 , 1874) peuvent être fiables pour indiquer la richesse de la faune de Lassime ou ne sont pas dignes de confiance. Selon Benoist , la faune de Lassime est originale, distincte de celle des niveaux équivalents connus en Aquitaine (la soi-disant «Sallomacienne» de Gironde [à Salles]; la «Helvétienne» des Pyrénées-Atlantiques [à Orthez]). Cela soulève de nombreuses questions. L'une d'elles concerne la citation de Benoist (1874 : p. 287) de l'occurrence de l'espèce récente " Modulus lenticularis (= Modulus module)" représentée par deux spécimens de sa collection (notre Pl. 1 , fig. 1-8) . Aucun autre spécimen de moduleétait connue du Serravallien du Bassin d'Aquitaine et le groupe d'espèces Modulus modulus était considéré comme exclusivement américain. Cependant, les spécimens de la collection Benoist de "Lassime" sont vraiment très proches du module module des Caraïbes (voir Pl. 1 , fig. 9-12). Parce que Benoist a également acquis une certaine (peut - être injustifiée) la réputation de faussaire, au moins dans le domaine de l' archéologie (Gendrey , 2015), le premier auteur (P. Lozouet) avait une hésitation de longue date avant de considérer que ces Modulus vraiment venu du Serravallien d'Aquitaine.
Aujourd'hui, la consultation d'une autre collection historique de Lassime (réalisée au XIXe siècle) conservée à l'Université de Bordeaux (UB) dissipe enfin toute ambiguïté car parmi les quelques lots de mollusques conservés, il y a un spécimen de Modulus identique à ceux du Benoist collection. Par conséquent, nous confirmons que le groupe d'espèces Modulus modulus est bien présent dans le Miocène moyen de l'Aquitaine. Bien que nous ne puissions pas affirmer que les listes de Benoist sont correctes, cette découverte conduit à croire que certains des enregistrements sont corrects. De plus, les mollusques de la collection UB présentent des sédiments de marnes sableuses gris foncé incrustés, comparables à ceux de Linder (1872: p. 472) description d'un affleurement sur la rive gauche de la rivière (en contrebas de la ferme de Cazenave, en amont de Lassime) qui alimentait de nombreux petits gastéropodes. Dans cet affleurement, les mollusques étaient nettement mieux conservés que ceux de la section classique actuellement encore visibles sur la rive droite du fleuve.
3. Modulidae dans la collection Grateloup
De 1827 à 1847, le médecin Jean-Pierre Sylvestre Grateloup (1782-1861) a décrit plusieurs centaines d'espèces de gastéropodes des époques oligocène et miocène du bassin aquitain et particulièrement du bassin de l'Adour. Une grande partie de sa collection d'espèces nouvelles a été illustrée par Grateloup dans son célèbre « Atlas Conchyliologique » et dans d'autres articles. Cependant, ses dessins sont dans certains cas excessivement caricaturés et les descriptions très minimales ou absentes. Cela entrave la reconnaissance de l'espèce. De plus, le Grateloup La collection a été soumise à de nombreuses tribulations et mouvements au fil du temps. Un des deux meubles abritant historiquement la collection, a eu un accident de transport avec de nombreux tiroirs chavirant, entraînant un mélange de coquilles. En conséquence, de nombreux spécimens sont trouvés sans leurs étiquettes respectives. Par conséquent, une interprétation personnelle est nécessaire lors du reclassement et de la révision de la collection (Cahuzac et al. , 2004). La collection est actuellement hébergée à la Faculté des Sciences de l'Université de Bordeaux.
Nos travaux préliminaires d'arrangement des Trochoïdées de la collection Grateloup nous ont conduit à sélectionner un spécimen problématique illustré par Grateloup (1845 , Pl. 13, fig. 16). Grateloup (1832) nomma ce spécimen Trochus laevigatus , malheureusement un homonyme junior de Trochus laevigatus Gmelin , 1791, et Sowerby , 1817. Par conséquent, Orbigny (1852) introduisit le nom de Trochus sublaevigatus en référence à [Pl. 1, non. 13, fig. 16] à Grateloup (1845). GrateloupLe dessin n'est pas très fidèle et la flèche de la coquille a été restaurée (Fig. 3 ). Cependant, le spécimen récupéré semble avoir une forme et une taille similaires à celles de Grateloup .
Notre observation montre que Trochus sublaevigatus n'est pas un Trochidae ou Trochoidea mais un membre des Modulidae, comme l'indique l'absence de structure nacrée et la columelle caractéristique se terminant par une dent columellaire. En fait, la forme trochoïdale et l'ouverture profondément lirate à l'intérieur permettent de classer cette espèce dans le genre américain Trochomodulus .
Figure 3 : Assemblage graphique des données historiques. A , montage de la plaque par Grateloup , 1845 (Pl.1, n ° 13) y compris Trochus laevigatus (Fig.16). B , étiquette (écriture de Benoist) indiquant le module lenticularis de la couche "La Sime" avec la bivalve Cardita jouanneti (verbatim: "Couche supérieure du niveau à C. jouanneti"). C , Les mêmes données sont rapportées dans latraduction de Benoist (1874 , p. 286): espèce rare à La Sime, de la zone " Cardita Jouanneti" .
4. Modulidés de l'Éocène et de l'Oligocène inférieur
Notre examen plus approfondi du matériel récemment collecté dans le bassin aquitain a révélé la présence de nouvelles espèces de l'Oligocène inférieur et de l'Eocène supérieur. L'espèce de l'Oligocène inférieur est la plus ancienne occurrence du genre Trochomodulus. L'espèce Priabonian, classée dans le genre Incisilabium , est l'une des plus anciennes espèces signalées chez les Modulidae. Nous décrivons ces espèces ci-dessous.
5. Description systématique
Famille Modulidae P. Fischer , 1884
La classification des Modulidae est basée pour le moment sur la morphologie générale de la coquille. L'ouverture, et en particulier la zone siphonale (Landau et al ., 2014 ; voir aussi Lozouet & Krygelmans , 2016 , Fig.2), sont considérées comme les principales caractéristiques diagnostiques. Les genres Modulus et Trochomodulus partagent la même dépression siphonale renforcée qui est bien délimitée par une crête siphonale. Dans le groupe module module , la dépression siphonale est particulièrement bien développée et en forme de cuillère. Landau et coll. (2014) divisent la famille des Modulidae en six genres. Parmi eux, Laevimodulus et Psammodulus sont exclusivement des genres d'Amérique tropicale fossile, avec des enregistrements dans le Miocène-Pliocène. Trochomodulus était jusqu'à présent également considéré comme un genre exclusivement tropical américain représenté de la fin du début du Miocène à nos jours, dans les Caraïbes et au Panama. De nouvelles données révélées ici montrent que le groupe est également présent dans les gisements oligocènes européens.
Localité. France, Bassin d'Aquitaine, Saucats "Lassime".
Âge. Miocène moyen (serravallien).
Étymologie. Dédié à É.-A. Benoist qui a identifié cette espèce pour la première fois et a collecté deux spécimens précieux.
Dimensions. Holotype: SL = 8,5 mm, SD = 10,8 mm, SWL = 7,2 mm; Paratype 1: SL = 7 mm, SD = 8 mm, SWL = 6,3 mm; Paratype 2: SL = 17 mm, SD = 18 mm, SWL = 9 mm.
La description. Coquille de taille moyenne, solide, angulaire, turbiniforme avec une grande forme pyramidale et une flèche déprimée. Protoconch inconnu. Téléoconque de plus de 3,5 spires séparées par une suture, profondément imprimée et ondulante. Sculpture sur la flèche avec 4 cordons en spirale, trois fins et un très grand au-dessus de la ligne de suture; onze côtes axiales obsolètes, irrégulières, ondulées, mal délimitées. Coquille entière couverte de nombreuses et fines lignes de croissance.
Le verticille du corps occupe 85% de la longueur totale de la coquille. Rampe de suture large, arrondie, épaule anguleuse. Sculpture de rampe de suture constituée de 10 à 11 nervures axiales obsolètes s'arrêtant à la périphérie et de 5 cordons primaires avec 1 cordon secondaire dans les inter-espaces; puis un cordon primaire très solide délimitant la rampe; sculpture de la base composée de 5-6 cordons primaires et 2-3 cordons secondaires sur le cou. Ombilic petit mais profond. Grande ouverture subquadrate avec une fine callosité sur la partie pariétale; canal siphonal avec encoche siphonale et dépression siphonale renforcée en forme de cuillère; canal anal non développé. Columelle profondément creusée au milieu, avec un callus développé et une forte dent columellaire abapicale. Lèvre extérieure, tranchante, prosocline, profondément lirée à l'intérieur, par sept lires.
Discussion. La similitude du module benoisti et du module module est frappante (Pl. 1 ). Nous avons comparé l'ancien avec des spécimens subfossiles de module Module (Pl 1. , Les figures 9-12.) Ont été recueillies hors île Martinique durant l'expédition Madibenthos (Martinique, Gros Islet, Stn AD618 1m, 14 ° 32.9'N - 61 ° 01.3'W). Le module module présente une grande variation (voir Lamy & Pointier , 2017 , Pl.39 , Fig.3a -f) et Landau et al. (2014, Figues. 2-8) soupçonnait un complexe d'espèces. La seule différence significative entre les espèces d'Aquitaine et des Caraïbes semble être les cordons spiralés plus solides des coquilles des Caraïbes. L'espèce commune européenne du Miocène précoce Modulus basteroti Benoist , 1874 (Pl. 3 , fig. 1-4) présente une forme avec une flèche plus haute et n'a pas la dépression siphonale renforcée en forme de cuillère observée dans le groupe d'espèces Module module . En fait, le placement générique de M. basteroti est discutable (Lozouet & Krygelmans , 2016) et ne peut être résolu sans reconsidérer la classification de la famille, mais il ne semble pas appartenir à laGroupe d'espèces de module module . Modulus basteroti est cité du Miocène moyen des Paratethys et de l'Italie. Sacco (1896 , Pl. 1, fig. 2-4) a décrit deux variétés de M. basteroti du "Colli Torinesi". Les deux ont une flèche plus haute et des côtes axiales plus proéminentes que M. benoisti .
Trochomodulus Landau , Vermeij et Reich , 2014
Espèce type: Modulus catenulatus Philippi , 1849 (par désignation originale). Localité / âge: Pacifique Est, récent.
Localité. France, Bassin d'Aquitaine, Gaas "Lagouarde".
Âge. Oligocène inférieur (rupélien).
Étymologie. Espèce connue uniquement de l'Oligocène inférieur (Stampian = Rupelian).
Dimensions. Holotype: SL = 15,5 mm, SD = 15 mm, SWL = 12 mm; Paratype 1: SL = 2,4 mm, SD = 2,3 mm, SWL = 1,8 mm (Landes, Orist).
Autre matériel examiné. 4 spécimens , Landes, Oriste.
Description (holotype). Coquille de taille moyenne, presque aussi haute que large, solide, trochiforme de forme pyramidale. Protoconch inconnu. Téléconch de plus de 5 spires faiblement concaves séparées par une suture imprimée. Sculpture sur flèche avec 4-6 cordons spiralés granulaires primaires, cordon abapical plus fort. Coquille entière couverte de nombreuses lignes de croissance fines. Le verticille du corps occupe 77% de la longueur totale de la coquille, angulaire à la périphérie; sculpture composée de 7 cordons granulaires primaires avec un cordon périphérique solide en forme de quille et 1 cordon secondaire dans les espaces intermédiaires; sculpture sur socle composée de 6 cordes. Ombilic presque fermé. Grande ouverture subquadrate avec une fine callosité sur la partie pariétale; canal siphonal avec encoche siphonale, canal anal et dépression siphonale non développés. Columelle creusée au milieu, avec une dent columellaire émoussée, cal non développé mais probablement usé.
Remarque. Paratype 1 (jeune spécimen) avec 4 cordons spiralés sur l'avant-dernière verticille. Cordon adapical avec de très gros granules. Le cordon adapical et le cordon abapical sont plus gros. Corde abapicale sur la dernière verticille presque lisse, formant une forte quille délimitant la base.
Discussion. Trochomodulus stampinicus se sépare facilement de T. sublaevigatus par ses cordons granuleux. Le même constat s'applique aux espèces américaines, dont aucune ne possède de tels cordons granuleux en spirale (Landau et al. , 2014).
1852 Trochus sublaevigatus d' Orbigny : no. 642, 42.
Matériel examiné. Oligocène supérieur, Chattien: lectotype désigné ici (SL = 16,2 mm, SD = 13 mm) (Pl. 2 , fig. 4-6), coll. Grateloup [UB-65-2-141] spécimen de Grateloup (1845 , Pl. 13, fig. 16), Landes, St-Paul-lès-Dax "faluns jaunes". Landes, St-Paul-lès-Dax ["Abesse, Château" 1 exemplaire (Pl. 2 , fig. 7-10), MNHN.FA71171; Lestrilles 5 spécimens, Pl. 2 , fig. 1-3, MNHN.FA71170].
Discussion. Trochomodulus paraguanensis Landau et al. , 2014 , du Miocène inférieur (Burdigalien) du Venezuela est de forme similaire à T. sublaevigatus ; cependant, les cordons spiralés sont moins nombreux et plus pointus chez T. paraguanensis (voir Landau et al. , 2014 , fig. 54-58).
Incisilabium Cossmann , 1918
Espèce type: Monodonta parisiensis Deshayes , 1832 (par désignation d'origine). Localité / âge: Bassin de Paris, Eocène moyen, Bartonien.
Localité. France, Bassin d'Aquitaine, Peyrehorade "Lasserre".
Âge. Éocène supérieur (Priabonien).
Étymologie. De la forme de la coquille.
Dimensions. Holotype: SL = 5,1 mm, SD = 4,2 mm, SWL = 12 mm; Paratype 1: SL = 4,4 mm, SD = 4,5 mm; Paratype 2: SL = 3,7 mm, SD = 3,7 mm; Paratype 3: spécimen juvénile.
Autre matériel examiné. 23 jeunes spécimens, Landes, Peyrehorade "Lasserre".
Description (holotype). Coquille de petite taille, presque aussi haute que large, solide, trochiforme de forme pyramidale. Protoconch de type larvaire planctotrophe, de 2,5 verticilles convexes, avec un petit noyau, bien démarqué de la téléoconque; Protoconch I lisse, Protoconch II avec filetages en spirale et riblets axiaux. Téléoconque (usée) de 4-5 spires, séparées par une fine suture imprimée. Spire faiblement concave avec 4-6 cordons spiralés primaires, cordon abapical plus fort. Le verticille du corps occupe 82% de la longueur totale de la coquille, rampe de suture large faiblement concave, épaule anguleuse. Rampe suturale large, délimitée par un cordon solide (angle d'épaule) et sculpté de 6-7 cordons, cordon adapical (le long de la suture) solide. Partie médiane du verticille (entre la base et la rampe de suture) avec 3 cordons; base arrondie, circonscrite par une corde solide, portant 11 à 12 cordes primaires; au total 22-24 cordons en spirale sur la verticille du corps. Ombilic fermé. Grande ouverture, sous-quadrillée, avec une fine callosité sur la partie pariétale; canal siphonal avec encoche siphonale faible; canal anal et dépression siphonale non développés. Columelle creusée au milieu, avec une dent columellaire émoussée. Lèvre extérieure avec un bord biseauté, prosocline, lisse à l'intérieur de l'ouverture.
Discussion. Incisilabium trochiformis est séparé de I. parisiensis (Pl. 3 , fig. 5-9) par ses cordons spiralés non granuleux. Il se distingue du Trochomodulus par la faiblesse de son encoche siphonale et de sa dent columellaire ainsi que par sa lèvre extérieure intérieure lisse.
6. Conclusions
Le module européen du Miocène moyen benoisti n. sp., qui appartient au groupe d' espèces des Caraïbes Modulus modulus , est un exemple frappant d'espèce fossile difficile à distinguer d'une espèce récente géographiquement et stratigraphiquement éloignée. Jusqu'à présent, Trochomodulus était considéré comme un genre exclusivement tropical américain, allant de la fin du début du Miocène à aujourd'hui dans les provinces tropicales des Caraïbes et du Pacifique (Landau et al. , 2014). Sa présence en Europe depuis au moins l'Oligocène inférieur était inattendue. En fait, Modulus benoisti , Trochomodulus stampinicus et Trochomodulus sublaevigatuspointent vers des échanges transatlantiques de taxons au cours de l'Oligocène et du Néogène mais aussi à un biais dans les archives fossiles. Les modules fossiles sont relativement rares au Cénozoïque et peuvent être facilement confondus avec les Trochoidea, comme en témoigne une mauvaise interprétation de Trochus sublaevigatus . En conséquence, le plus ancien membre non ambigu des Modulidae a été signalé à l'Eocène moyen de l'Europe avec le genre Incisilabium Cossmann , 1918 (Lozouet & Krygelmans , 2016 , Fig.2F ; Cossmann & Pissarro , 1907 , Pl.3 , Fig.22-1) anciennement classée dans les Trochidae. C'est la raison pour laquelle Landau et al. (2014) ont négligé cette référence et considéré que les Modulidae les plus anciens connus provenaient de l'Éocène supérieur / Oligocène inférieur américain. L'affirmation de ces auteurs selon laquelle «les archives fossiles indiquent fortement une origine tropicale américaine pour les Modulidae» n'est donc plus valable.
La phylogénie moléculaire par Strong et al. (2011) montre que les Modulidae / Scaliolidae sont très proches des Potamididae et les place à la base d'un clade réunissant les Paludomidae et Thiaridae d'eau douce. Cette phylogénie implique une divergence très précoce pour les Modulidae. Enfin, ces nouvelles données confirment en outre que l'histoire paléobiogéographique des Modulidae est plus complexe que prévu et que l'origine de la famille est probablement plus ancienne que l'époque éocène. On peut donc s'attendre à la découverte d'espèces antérieures de Modulidae dans le futur.
Remerciements
Nous remercions Philippe Rocher (RNG Saucats-La Brède) Laurent Londeix (UB, Bordeaux) qui a fourni des informations sur le RNG, et Jean-Michel Pacaud (MNHN-Paris) pour le prêt de matériel. Merci à Geerat Vermeij (Davis, USA) et Jean-Claude Plaziat (Paris) pour leurs commentaires. Merci également à Claudia Ratti (MNHN, Paris) pour avoir perfectionné l'anglais du manuscrit.
Références bibliographiques
Benoist É.-A. (1873).- Catalogue synonymique et raisonné des testacés fossiles recueillis dans les faluns miocènes des communes de La Brède et de Saucats.- Actes de la Société Linnéenne de Bordeaux, tome 29, p. 5-78.
Benoist É.-A. (1874).- Catalogue synonymique et raisonné des testacés fossiles recueillis dans les faluns miocènes des communes de La Brède et de Saucats.- Actes de la Société Linnéenne de Bordeaux, tome 29, p. 265-460.
Bouchet P. (2018).- MolluscaBase. Indomodulus Landau, Vermeij & Reich, 2014. URL: http://www.molluscabase.org/aphia.php?p=taxdetails&id=817008 [last accessed on 2018-10-04].
Cahuzac B. (2005).- Découverte de Pecten praebenedictus (Bivalvia, Pectinidae) dans le Serravallien de Saucats (SW France) ; aperçu sur les Pectinidae de ce site.- Bulletin de la Société Linnéenne de Bordeaux (nouv. série), tome 140, no. 33 (1), p. 21-42.
Cahuzac B. & Cluzaud A. (1999a).- Bilan scientifique et synthèse des données bibliographiques sur le Miocène moyen (Serravallien) de Saucats (Gironde, Bassin d'Aquitaine).- Bulletin de la Société Linnéenne de Bordeaux, vol. 27, no. 2, p. 95-111.
Cahuzac B. & Cluzaud A. (1999b).- Bilan scientifique et synthèse des données bibliographiques sur le Miocène moyen (Serravallien) de Saucats (Gironde, Bassin d'Aquitaine). (suite et fin).- Bulletin de la Société Linnéenne de Bordeaux, vol. 27, no. 3, p. 113-133.
Cahuzac B., Turpin L. & Bonhomme B. (1997).- Sr isotope record in the area of the Lower Miocene historical stratotypes of the Aquitaine Basin (France). In: Montanari A., Odin G.S. & Coccioni R. (eds.), Miocene Stratigraphy - an integrated approach.- Developments in Palaeontology and Stratigraphy, vol. 15, chapter A 4, p. 33-56.
Cahuzac B., Lesport J.-F. & Lagarde L. (2004).- Révision des Cancellariidae (Mollusca, Gastropoda) décrites par Grateloup (1827-1847) dans le Miocène des Landes (SW France).- Geodiversitas, Paris, vol. 26, no. 2, p. 207-261.
Cossmann M. & Peyrot A. (1909-1924).- Conchologie néogénique de l'Aquitaine.- Actes de la Société Linnéenne de Bordeaux, vol. LXIII (2), 1909, p. 72-144 (Pls. I-IV); vol. LXIII (3), 1909, p. 145-232 (Pls. V-VII); vol. LXIII (4), 1909, p. 233-293; t. LXIV (4), 1910, p. 235-288; t. LXIV (5), 1910, p. 289-400; t. LXIV (6), 1911, p. 401-445 (Pls. VIII-XVIII); t. LXV (2), 1911, p. 51-98; t. LXV (3), 1912, p. 99-178; t. LXV (4), 1912, p. 179-333 (Pls. XIX-XXVIII); t. LXVI (2), 1912, p. 121-168; t. LXVI (3), 1912, p. 169-232; t. LXVI (4), 1913, p. 233-324 (Pls. I-X); t. LXVIII (1), 1914, p. 5-96; t. LXVIII (2), 1914, p. 97-210 (Pls. XI-XXII); t. LXVIII (3), 1914, p. 361-435 (Pls. XXIII-XXVI); t. LXIX (3), 1917, p. 158-284; t. LXIX (4), 1917, p. 285-365; t. LXX (1), 1918, p. 5-100; t. LXX (2), 1918, p. 101-180 (Pls. I-X); t. LXX (3), 1919, p. 181-356; t. LXX (4), 1919, p. 357-491 (Pls. XI-XVII); t. LXXIII, 1922, p. 5-321 (Pls. I-VII); t. LXXIV (3), 1923, p. 257-342; t. LXXV (2), 1924, p. 71-144; t. LXXV (3), 1924, p. 193-318 (Pls. I-VII).
Cossmann M. & Pissarro G. (1907).- Iconographie complète des coquilles fossiles de l'Éocène des environs de Paris.- Hermann, Paris, Tome II, pars - In-4º, Pls. I-IX. URL: https://www.biodiversitylibrary.org/bibliography/52315#/
Gendry D. (2015).- Deux paléontologues berrichons: André Émile Benoist (1845-1904) et l'abbé Alexis Delaunay (1867-1946), et leur contribution à l'étude du Bathonien de Saint-Gaultier (Indre, France).- Symbioses (nouvelle série), vol. 33, p. 1-16.
Grateloup J.-P.S. (1832).- Tableau (suite du) des coquilles fossiles qu'on rencontre dans les terrains calcaires tertiaires (faluns) des environs de Dax, dans le département des Landes.- 4ème article.- Actes de la Société Linnéenne de Bordeaux, tome 5(3), no. 27, p. 132-171.
Grateloup J.-P.S. (1845).- Conchyliologie fossile des terrains tertiaires du Bassin de l'Adour (environs de Dax). 1, Univalves. Atlas.- Lafargue, Bordeaux, 45 planches et leurs textes explicatifs (1, 3, 5-10, 12-48).
Lamy D. & Pointier J.-P. (2017).- Marine and freshwater molluscs of the French Caribbean.- PLB Editions, Gosier, vol. 1, 388 p.
Landau B., Vermeij G.J. & Reich S. (2014).- Classification of the Modulidae (Caenogastropoda, Cerithioidea), with new genera and new fossil species from the Neogene of tropical America and Indonesia.- Basteria, Bilthoven, vol. 78, no. 1-3, p. 1-29.
Linder O. (1872).- Des dépôts lacustres du vallon de Saucats.- Actes de la Société Linnéenne de Bordeaux (3ème série), vol. 27 (1870-1872), p. 451-525.
Londeix L. & Jan Du Chêne R. (1998).- Burdigalian dinocyst stratigraphy of the stratotypic area (Bordeaux, France).- Geobios, Villeurbanne, vol. 30, no. 2, p. 283-294.
Lozouet P. & Krygelmans A. (2016).- A new species of Indo-Pacific Modulidae (Mollusca: Caenogastropoda).- Zootaxa, Auckland, no. 4103 (2), p. 195-200.
Mayer K. (1858).- Versuch einer neuen Klassifikation der Tertiär-Gebilde Europa's.- Verhandlungen der allgemeinen Schweizerischen Gesellschaft für die gesammten Naturwissenschaften, Basel, p. 70-71, 165-199.
Orbigny A. d' (1852).- Prodrome de paléontologie stratigraphique universelle des animaux mollusques et rayonnés.- vol. 3, 191 p.
Parize O., Mulder T., Cahuzac C., Fiet N., Londeix L. & Rubino J.-L. (2008).- Sedimentology and sequence stratigraphy of Aquitanian and Burdigalian stratotypes in the Bordeaux area (southwestern France).- C.R. Geoscience, Paris, vol. 340, p. 390-399.
Peyrot A. (1925-1935).- Conchologie néogénique de l'Aquitaine.- Actes de la Société Linnéenne de Bordeaux, t. LXXVII (2), 1925, p. 51-194; t. LXXVIII, 1927, p. 199-256 (Pls. I-IV); t. LXXIX suppl., 1928, p. 5-264 (Pls. V-XIV); t. LXXXII (2), 1931, p. 73-126; t. LXXXIII, 1931, p. 5-116 (Pls. I-X); t. LXXXIV (1), 1932, p. 5-128; t. LXXXIV (2), 1933, p. 129-288 (Pls. XI-XVIII); t. LXXXV (1), 1933, p. 5-71; t. LXXXVI (2), 1935, p. 257-352.
Sacco F. (1896).- I molluschi dei terreni terziarii del Piemonte e della Liguria. Parte 21.- Carlo Clausen, Torino, 60 p. (4 Pls.).
Strong E.E., Colgan D.J., Healy J.M., Lydeard C., Ponder W.F. & Glaubrecht M. (2011).- Phylogeny of the gastropod superfamily Cerithioidea using morphology and molecules.- Zoological Journal of the Linnean Society, London, vol. 162, p. 43-89.