(traduction post-éditée par N. Bacaër, suggestions d'amélioration : nicolas.bacaer@ird.fr)
Résumé
Il existe peu d’information à propos des capacités cognitives des lamantins et comment elles peuvent permettre d’améliorer le bien-être de ces animaux en captivité. Notre étude analyse comment la présence de nouveaux stimuli et leur association avec de la nourriture peut changer le comportement d’un lamantin antillais nommé Daniel. Premièrement Daniel a été observé en absence de stimuli puis ensuite en présence de quatre formes géométriques différentes. Dans un troisième temps Daniel fût entrainé à manger en association avec une forme, le carré. Dans un quatrième temps, les quatre formes lui sont proposées mais sans nourriture associée. Les comportements différents et les interactions du lamantin avec le carré augmentent considérablement après cet aprentissage. L’expèrience est retentée 3 mois et 12 mois plus tard, sans renforcement, et Daniel continue de choisir le carré avec des comportements particuliers liés à cette forme. Cette étude permet de montrer de manière formelle la capacité des lamantins à associer la possibilité de nourriture avec des formes visuelles, et aussi la possibilité d’augmenter son activité avec des dispositifs environnementaux et occupationnels. Ces résultats offrent de nouvelles perspectives pour l’étude du comportement des lamantins, en particulier liées à la cognition, à leur gestion et à leur bien-être en captivité.
1. Introduction
L'apprentissage est courant chez les animaux, des petits invertébrés aux mammifères et joue un rôle important en terme d'adaptation [ 1 ]. L'apprentissage associatif qui consiste en l'association de deux éléments différents [ 2 ] est particulièrement pertinent pour l'étude de la cognition animale [ 3 ].
La recherche sur la cognition chez les mammifères marins est bien documentée, en particulier pour les cétacés, avec des études principalement axées sur les grands dauphins et les pinnipèdes, principalement les otaries de Californie [ 4 , 5 ]. Au cours des dernières années, la recherche sur la cognition animale a souvent été associée à des études sur le bien-être animal, comme le suggèrent plusieurs auteurs [ 4 , 6 , 7 ]. L'enrichissement environnemental, associé au concept de bien-être chez les animaux, a d'abord consisté en des améliorations de l'environnement animal captif [ 8 ]. Ce concept relativement nouveau a été bien étudié chez les mammifères aquatiques, en particulier les dauphins [ 9]. Plusieurs études suggèrent que l'enrichissement de l'environnement pour le bien-être animal permet à l'animal d'exprimer des comportements et d'utiliser des capacités, telles que la cognition [ 7 ]. La stimulation des capacités cognitives chez les animaux, par l'enrichissement en captivité, est une technique qui peut augmenter le bien-être animal et diminuer, voire éliminer, les comportements stéréotypés chez un grand nombre d'animaux [ 10 ]. L'enrichissement sensoriel par l'enrichissement environnemental ou professionnel chez les animaux peut réduire ou éliminer les comportements stéréotypés et augmenter le bien-être qui peut être mesuré par l'amélioration des modèles de comportement [ 11]. Concernant les capacités cognitives, de nombreuses études ont démontré que les animaux, les invertébrés ou les vertébrés possèdent des capacités d'apprentissage qui sont utilisées lorsqu'ils doivent échapper à des prédateurs ou à un danger [ 12 ], ou lorsqu'ils se nourrissent pour augmenter leurs chances de succès [ 13 , 14 ]. Ces capacités sont bien étudiées chez les mammifères aquatiques, comme les dauphins ou les épaulards [ 5 ]. L'apprentissage est généralement lié à la capacité des animaux à faire la distinction entre les signaux, en particulier visuels, comme cela a été démontré chez les insectes [ 15 ], les oiseaux [ 16 ] ou les mammifères [ 17 , 18 ]. Chez les mammifères aquatiques, la discrimination entre les signaux est bien étudiée dans un contexte social, par exemple fTursiops truncatus Montagu 1821 (Cetaceae, Delphinidae) qui utilise des contours de sifflet pour discriminer les conspécifiques [ 19 ]. Bien que les dauphins soient considérés comme des spécialistes de l'acoustique, les indices visuels peuvent également jouer un rôle important et ont été utilisés dans plusieurs études sur la cognition des dauphins. Par exemple, Kilian et al. [ 20 ] ont étudié la capacité d'une femelle dauphin à discriminer les stimuli visuels différant en nombre. Une autre étude montre qu'une femelle béluga était également capable de discriminer visuellement les quantités de nourriture [ 21 ].
Les études comportementales sur le lamantin des Antilles ( Trichechus manatus manatus , Linnaeus 1785; Sirenia, Trichechidae) sont rares, malgré leur importance pour la conservation et leur présence dans plusieurs zoos et parcs. Hartman [ 22 ] a réalisé la première étude à grande échelle du comportement des lamantins et a déclaré que les lamantins n'ont pas besoin de modèles de comportement complexes en raison du manque de prédateurs, de l'abondance de nourriture et des températures stables de l'eau où ils vivent. Koelsch [ 23 ] a suggéré un niveau de sociabilité plus élevé que celui décrit par Hartman [ 22 ] dans Bahía de Sarasota (Floride), tandis que Harper et Schulte [ 24 ] ont observé que les interactions sociales se produisaient uniquement entre les femmes. La dernière étude de Hénautet coll. [ 25 ] ont rapporté qu'en captivité, des interactions sociales se produisaient également entre mâles et femelles et en particulier entre un veau et les divers membres du groupe des lamantins, quels que soient le sexe et l'âge. Un seul travail récent s'est concentré sur le bien-être des lamantins, décrivant leurs comportements stéréotypés [ 26 ]. Comme observé chez plusieurs mammifères marins, le «comportement de nage en cercle» semble être la caractéristique comportementale typique du malaise du lamantin en captivité [ 26 ]. À ce jour, aucune étude n'a adopté une approche comportementale de la perception visuelle et des capacités cognitives du lamantin, malgré l'importance que ce type d'étude peut avoir sur leur gestion et en particulier sur le bien-être des animaux captifs et leur réintroduction réussie dans la nature.
La vision chez les siréniens est considérée comme limitée par rapport aux autres mammifères aquatiques [ 27 ]. Cependant, Hartman [ 21 ] a rapporté que la vision peut être importante pour les lamantins et selon Reynolds et Oddel [ 28 ] les yeux des lamantins sont capables de voir en plein jour et dans la pénombre. Reep et Bonde [ 27 ] ont décrit que les lamantins ont une vision dichromatique observée chez les pinnipèdes, avec la détection des bleus et des verts. Les lamantins ont des poils tactiles qui offrent une bonne discrimination (par exemple pour explorer la végétation) et sont également capables d'utiliser leurs poils faciaux pour détecter toutes les directions de leur environnement lorsqu'ils nagent dans un motif circulaire [ 29]. Bien que la taille du cerveau du lamantin soit petite par rapport à la taille du corps, le cortex cérébral est similaire à celui des mammifères à grand cerveau, ce qui peut indiquer des capacités cognitives considérables [ 30 ].
Peu d'études se sont concentrées sur le bien-être des lamantins en captivité. Ces mammifères sont généralement captifs dans un espace réduit, surtout par rapport à leur milieu naturel, et la plupart du temps, ils peuvent être observés nageant autour de l'aquarium [ 26 ]. Ce comportement stéréotypé chez les lamantins semble associé à un répertoire comportemental réduit (Hénaut, observation personnelle). Ce répertoire réduit peut aussi être la conséquence d'une mauvaise gestion des lamantins captifs, avec un manque d'activités stimulantes et peu d'interactions sociales avec les congénères ou les humains, même en tenant compte des soins vétérinaires [ 31 ], ou lorsqu'ils sont nourris par les touristes [ 25]. En outre, le faible nombre d'études liées au bien-être des lamantins et à leurs capacités cognitives garantit que les nouvelles informations sur le comportement des lamantins, en particulier si elles sont axées sur la cognition et le bien-être, sont non seulement fascinantes en termes de contribution scientifique unique, mais également précieuses pour leur gestion en captivité et pour réussir les réintroductions.
Les études comportementales des lamantins se produisent généralement avec des individus captifs dans des centres zoologiques ou touristiques. Les contraintes de captivité associées ne fournissent pas les conditions appropriées pour les observations comportementales, en particulier pour les études cognitives, car la forme prédominante de gestion limite les possibilités d'utiliser des dispositifs ou d'introduire des objets dans leur bassin et le public dérange souvent les animaux individuels. De plus, les lamantins en captivité vivent généralement en groupes avec interaction entre les animaux individuels [ 25 ]. Il est également difficile d'accéder à un grand nombre de lamantins pour une observation individuelle sans possibilité de perturbation sociale, en particulier sur l'apprentissage social, effet que l'on attendrait pour de nombreuses espèces animales [ 32 ].
Heureusement, nous avons pu étudier le comportement d'un seul lamantin, appelé Daniel, élevé en semi-captivité. Le but de notre travail est d'étudier les capacités d'apprentissage liées à l'association alimentaire avec les repères visuels et la capacité de mémoire du lamantin Daniel.
2. Matériel et méthodes
2.1. Animal et logement
Les observations ont été menées dans le «Centro de Atención y Rehabilitación de Mamíferos Acuáticos» (CARMA) (Centre pour l'attention et la réhabilitation des mammifères aquatiques) situé dans une lagune (Laguna Guerrero,) près de Chetumal, dans l'état de Quintana Roo, au sud -Est du Mexique. Dans ce centre, un lamantin appelé Daniel, un garçon de 7 ans, vit dans des conditions «semi-libres». Daniel, veau orphelin sauvé à sa naissance récente, vit dans un espace clos où la nourriture est fournie, et peut régulièrement sortir dans le lagon à volonté, généralement l'après-midi. Pratiquement tous les jours, Daniel revient dans la région avant la tombée de la nuit. La zone est rectangulaire (17,30 m × 15 m) entourée d'une enceinte en bois et treillis métallique. La profondeur de l'eau était d'environ trois mètres. L'eau est suffisamment claire pour permettre une bonne observation de Daniel.
2.2. Matériel
Nous avons utilisé quatre objets géométriques (carré, triangle équilatéral, disque et hexagone) en acrylique blanc (Figure 2 ). Le carré et le triangle avaient des côtés de 50 cm tandis que le disque et l'hexagone avaient un diamètre de 50 cm; par conséquent, tous les objets étaient de taille similaire. Chaque forme avait un tube en PVC blanc de 2,5 cm le long de son bord inférieur afin qu'il puisse être immergé verticalement. La forme a été suspendue à l'enceinte avec une corde de sorte que la partie supérieure soit à seulement 1 cm sous la surface de l'eau (voir Vidéos supplémentaires, 1, 2, 3 et 4).
Lorsque les observations ont été faites à l'extérieur de la zone fermée, nous avons fait un petit trou de 1 cm de diamètre au centre de chaque forme afin que la nourriture puisse être attachée à l'aide d'une ligne de pêche. La forme était suspendue à un tube à air fixé sur une matraque en bois martelée dans le fond de la lagune.
La laitue Cos ( Lactuca sativa L. var. Longifolia) était utilisée comme nourriture car les observations précédentes suggèrent que c'était sa préférée.
Les formes étaient toujours espacées d'au moins 2,5 m et leurs positions changées après chaque observation, en utilisant toute la zone fermée. L'observateur était toujours la même personne et changeait de position pour être toujours à distance des formes pour éviter de déranger Daniel. En effet, Daniel est habitué à la présence d'un observateur et n'a présenté aucun comportement particulier orienté vers l'observateur lors de nos expériences après au moins une journée de présence. Nous avons enregistré des vidéos de Daniel pour illustrer clairement les comportements.
2.3. Procédure expérimentale et tâche d'apprentissage associatif
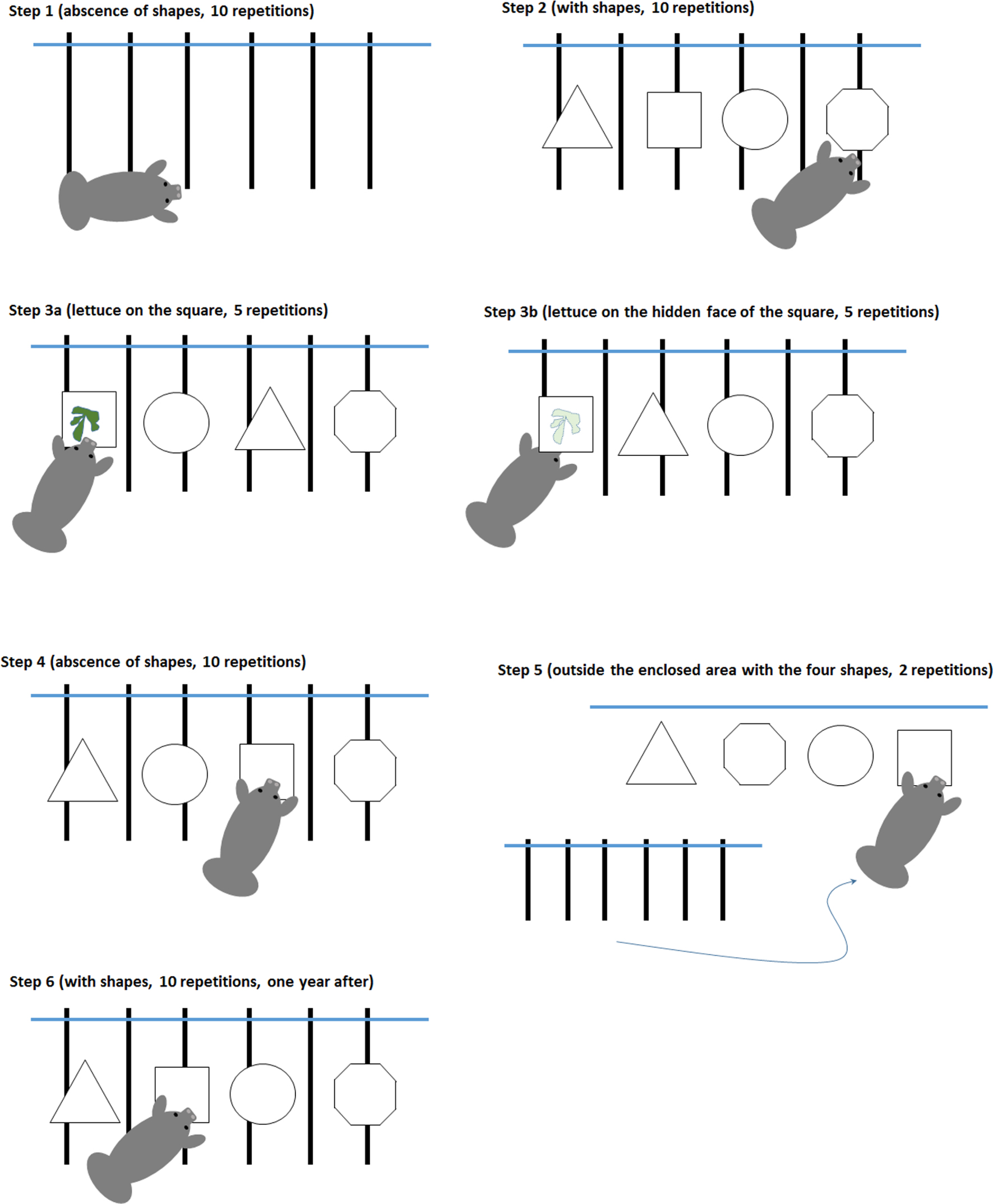
Figure 1. Les six étapes de la procédure expérimentale.
Figure 2.
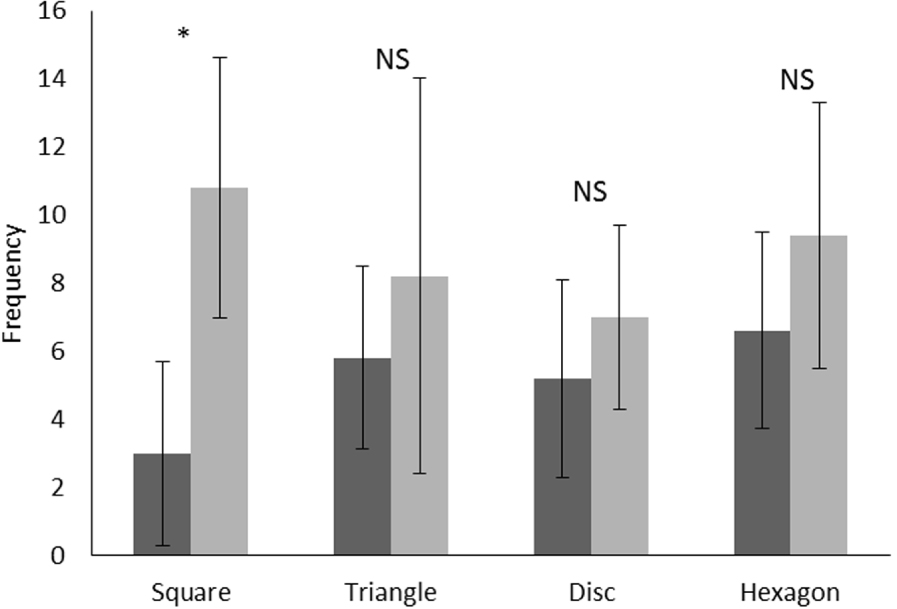
Erreur moyenne et standard de fréquence pour le nombre de chemins parcourus devant chaque forme par le lamantin, avant (étape 2, en noir) et après (étape 4, en gris) entraînement. Test de Wilcoxon, *: P <0,05, NS: non significatif.
Les observations se sont déroulées en six étapes (figure 1 ).
Étape 1. Nous avons observé le comportement de Daniel en l'absence de formes, 10 répétitions ont été effectuées.
Étape 2. Nous avons observé le comportement de Daniel en présence des quatre formes géométriques, immergées comme décrit précédemment, et sans nourriture (10 répétitions).
Étape 3 dite «étape d'apprentissage» qui consiste à sélectionner la forme la moins préférée par Daniel lors de l'étape 2 (le carré) puis à y attacher de la nourriture.
Étape 3a. Cinq répétitions ont été effectuées avec la laitue sur la face ouverte de la forme face au centre de la zone (visible par Daniel).
Étape 3b. 5 répétitions avec la nourriture sur la face cachée de la figurine.
Étape 4. Nous avons observé le comportement de Daniel, encore une fois avec les quatre formes, comme à l'étape 2, mais sans nourriture (10 répétitions).
Étape 5. Nous présentons les quatre formes à l'extérieur de la zone fermée et ouvrons l'enceinte pour que Daniel puisse partir (2 répétitions).
Étape 6. Nous avons observé le comportement de Daniel devant les quatre formes sans nourriture dans la zone fermée, 3 mois (1 observation) et 12 mois (1 observation) après «l'étape d'entraînement» et sans renfort ni accès aux formes pendant ce laps de temps .
Toutes les observations ont eu lieu de 8 h à 14 h, car les observations préliminaires indiquaient que c'était la période d'activité maximale de Daniel. Pour chaque répétition, nous avons observé l'animal pendant 30 min avec un maximum de cinq répétitions par jour. Nous avons enregistré tous les événements comportementaux pendant la période d'observation. Nous avons divisé la zone entourée par l'enceinte en quadrants virtuels (1 m 2 chacun) où nous avons noté sur le papier schématique le chemin de Daniel (ou «nager par» lorsque Daniel a traversé un quadrant), le comportement et le temps passé dans chaque quadrant.
2.4. Données et analyse
Lors de l'étape 1, sans aucune forme, nous avons noté le nombre de chemins parcourus par Daniel nageant et traversant un quadrant ainsi que le temps passé dans chaque quadrant à l'intérieur de l'enceinte où les formes seraient submergées lors de l'étape suivante. Nous avons considéré que pour toutes les étapes, à l'exception de la première (sans formes), une visite de Daniel était enregistrée lorsqu'il traversait un quadrant avec sa tête à au moins 50 cm de la forme présente dans le quadrant. Quand Daniel est resté dans un quadrant sans bouger, sa tête clairement orientée vers une forme comme s'il la regardait, nous avons enregistré que Daniel «regardait la figure». Lors de l'étape 1, nous avons enregistré le nombre de chemins dans des quadrants sans formes, où ils seraient ensuite installés dans les étapes supplémentaires. Nous avons également enregistré le temps passé par Daniel à traverser ces quadrants. La fréquence des visites et le comportement «regarder la forme» ont été enregistrés pour l'étape 2 (formes sans nourriture avant l'entraînement), l'étape 4 (formes sans nourriture après l'entraînement) et l'étape 5 (formes sans nourriture, après l'entraînement mais en dehors de la zone fermée ). Tous les autres comportements, y compris les comportements inattendus, ont été décrits dans les résultats. Lors de la «phase d'entraînement» (étapes 3a et 3b), nous avons enregistré le temps nécessaire pour «atteindre le chiffre», «commencer à manger la laitue» une fois le chiffre atteint, et «finir de consommer la laitue» jusqu'à ce que la laitue soit complètement mangée . Lorsque les formes étaient présentées à Daniel 3 et 12 mois après l'étape de formation (étape 6), seule l'interaction de Daniel avec les formes et les comportements associés à ces interactions était enregistrée. étape 4 (formes sans nourriture après l'entraînement) et étape 5 (formes sans nourriture, après l'entraînement mais en dehors de la zone fermée). Tous les autres comportements, y compris les comportements inattendus, ont été décrits dans les résultats. Lors de la «phase d'entraînement» (étapes 3a et 3b), nous avons enregistré le temps nécessaire pour «atteindre le chiffre», «commencer à manger la laitue» une fois le chiffre atteint, et «finir de consommer la laitue» jusqu'à ce que la laitue soit complètement mangée . Lorsque les formes ont été présentées à Daniel 3 et 12 mois après l'étape de formation (étape 6), seule l'interaction de Daniel avec les formes et les comportements associés à ces interactions a été enregistrée. étape 4 (formes sans nourriture après l'entraînement) et étape 5 (formes sans nourriture, après l'entraînement mais en dehors de la zone fermée). Tous les autres comportements, y compris les comportements inattendus, ont été décrits dans les résultats. Lors de la «phase d'entraînement» (étapes 3a et 3b), nous avons enregistré le temps nécessaire pour «atteindre le chiffre», «commencer à manger la laitue» une fois le chiffre atteint, et «finir de consommer la laitue» jusqu'à ce que la laitue soit complètement mangée . Lorsque les formes étaient présentées à Daniel 3 et 12 mois après l'étape de formation (étape 6), seule l'interaction de Daniel avec les formes et les comportements associés à ces interactions était enregistrée. Lors de la «phase d'entraînement» (étapes 3a et 3b), nous avons enregistré le temps nécessaire pour «atteindre le chiffre», «commencer à manger la laitue» une fois le chiffre atteint, et «finir de consommer la laitue» jusqu'à ce que la laitue soit complètement mangée . Lorsque les formes étaient présentées à Daniel 3 et 12 mois après l'étape de formation (étape 6), seule l'interaction de Daniel avec les formes et les comportements associés à ces interactions était enregistrée. Lors de la «phase d'entraînement» (étapes 3a et 3b), nous avons enregistré le temps nécessaire pour «atteindre le chiffre», «commencer à manger la laitue» une fois le chiffre atteint, et «finir de consommer la laitue» jusqu'à ce que la laitue soit complètement mangée . Lorsque les formes étaient présentées à Daniel 3 et 12 mois après l'étape de formation (étape 6), seule l'interaction de Daniel avec les formes et les comportements associés à ces interactions était enregistrée.
Pour déterminer si la tâche d'apprentissage associatif fonctionnait, nous avons comparé à l'aide d'un test de Friedman, le nombre de chemins, de visites, de comportements «regarder la forme» et le temps passé dans chaque quadrant pour les quatre formes lors de l'étape 2 (avant l'entraînement) et étape 4 (après l'entraînement). Nous avons comparé le temps passé par Daniel dans les quadrants avec une forme (étape 2) et le quadrant correspondant sans la forme (étape 1) à l'aide d'un test de paires appariées de Wilcoxon. Le même test a été utilisé pour comparer le nombre de visites pour ces deux étapes. Le temps nécessaire pour «atteindre la figure» avec la laitue, pour «commencer à manger» et «finir de consommer la laitue» pour la laitue devant la figure et la laitue sur la face cachée de la figure (étape 3a et 3b) ont été comparés en utilisant un test de paires appariées de Wilcoxon. Le nombre de chemins, de visites, Les comportements «regarder la forme» et le temps passé devant chaque forme pour l'étape 2 (avant l'entraînement) ont été comparés à l'étape 4 (après l'entraînement) pour chaque forme à l'aide d'un test de paires appariées de Wilcoxon. Tous les tests ont été effectués en utilisantStatistiques 7.0.
Nous avons également enregistré et décrit les comportements d'événements de courte durée pour les quatre premières étapes. Il était impossible d'observer la totalité des comportements lors de la cinquième étape car la distance de Daniel rendait l'observation difficile. L'occurrence de comportements connus et l'apparition de nouveaux ont été comparés pour les étapes 1 à 4. Ces résultats descriptifs nous aideront à comprendre si et comment la présence des formes et la formation ont pu affecter les activités de Daniel. Pour l'étape 6, (observation après 3 mois et 12 mois) nous avons seulement décrit la sélection de certaines formes et comportements spécifiques. Des extraits vidéo d'essais exemplaires sont inclus dans le matériel supplémentaire.
3. Résultats
3.1. Comportement de Daniel sans et avec les formes (étapes 1 et 2)
Le comportement de Daniel au cours de l'étape 1 consistait essentiellement en des comportements de mouvement (tableau 1 ) principalement en nageant et en respirant autour de la zone fermée. À l'occasion, Daniel affichait des comportements de «rotation» et de «sortie». Lorsque les formes étaient présentes dans la zone, l'activité de Daniel a radicalement changé avec beaucoup plus de mouvements que lors de l'étape 1, et une série de nouveaux comportements dirigés vers les formes: «regarder la figure», «étreindre la figure» (Vidéo supplémentaire 1), «touchez la figurine avec son visage», «touchez la figurine avec un flipper» et «jouez avec la figurine»; cependant, le comportement de «sortie de tête» a disparu au cours de l'étape 2 (tableau 1 ). Le comportement «toucher la figure avec un flipper» a démontré un intérêt manifeste pour les nouveaux objets.
Tableau 1.
Description et codes de tous les comportements décrits dans cette étude
Le nombre de chemins était statistiquement plus élevé (tableau 2 ) dans le quadrant avec un triangle (étape 2) que dans le quadrant correspondant sans le triangle (étape 1). Plus de chemins ont été observés dans le quadrant avec l'hexagone ou le disque (étape 2) que dans les mêmes quadrants sans ces formes (étape 1) (tableau 2 ); cependant, ces différences n'étaient pas significatives. Dans la première étape, seuls les quadrants avec le carré ont montré une tendance à moins de visites par rapport aux quadrants correspondants sans le carré.
Tableau 2.
Erreur moyenne et standard des numéros de chemin parcourus par le lamantin dans les quadrants sans forme (étape 1) et le même quadrant avec une forme (étape 2), pour les quatre formes géométriques testées
Daniel a passé beaucoup plus de temps dans le quadrant avec l'hexagone (étape 2) que dans le quadrant correspondant sans l'hexagone pendant l'étape 1 (tableau 3 ). Ce n'est que dans le cas de l'hexagone que Daniel a passé beaucoup plus de temps dans les quadrants avec la figure que sans. Le temps passé dans les quadrants avec le triangle ou le disque avait tendance à être plus important que dans ceux sans ces formes, tandis que le temps passé dans le même quadrant avec le carré et sans le carré était identique. Le nombre de chemins empruntés par Daniel devant les formes (figure 2 ) et le nombre de visites (figure 3 ) ne présentaient pas de différences significatives entre les formes lors de la deuxième étape (test de Friedman, df = 3, F = 2,74, p= 0,43 et df = 3, F = 5,90, p = 0,11 respectivement).
Figure 3.
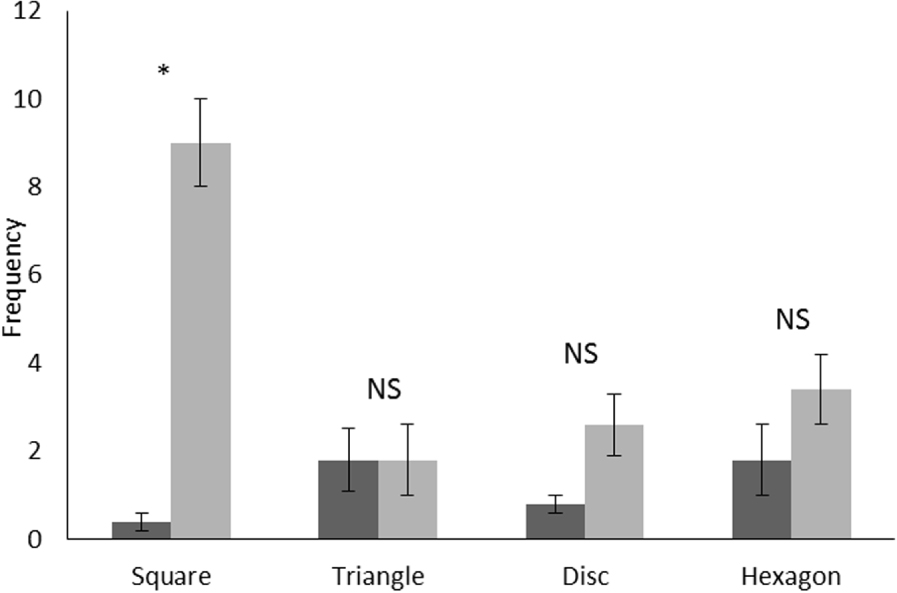
Erreur moyenne et standard de fréquence des visites effectuées par le lamantin avant (étape 2, en noir) et après (étape 4, en gris) l'entraînement. Test de Wilcoxon: NS: non significatif, *: P <0,05.
Tableau 3.
Temps moyen (min) passé par le lamantin dans des quadrants sans forme (étape 1) ou dans le quadrant correspondant avec une forme (étape 2), pour les quatre formes géométriques testées
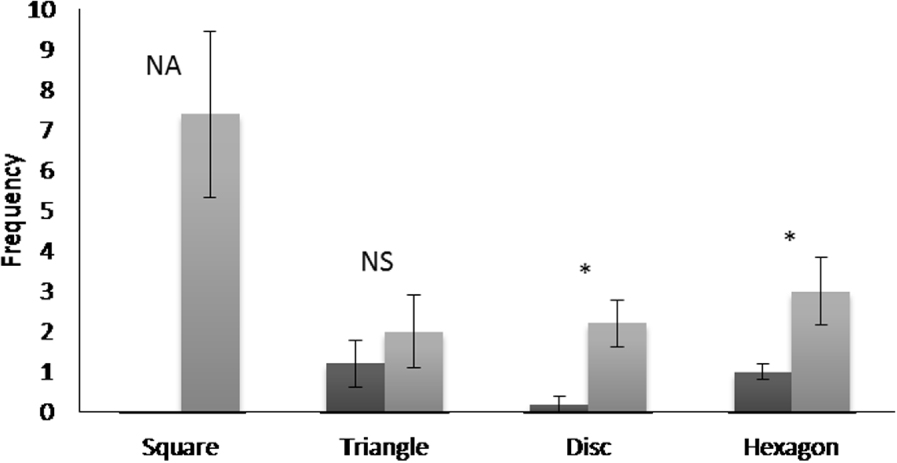
Le comportement «regarder la figure» n'a été observé que pour le triangle, le disque et l'hexagone lors de l'étape 2 (figure 4 ). Lors de la comparaison des fréquences de ce comportement, entre le triangle, le disque et l'hexagone à l'étape 2, aucune différence significative n'a été révélée (test de Friedman, df = 2, F = 4,90, p = 0,08). Daniel n'a jamais touché le carré mais a fréquemment touché les autres formes, en particulier l'hexagone qui était sujet à «étreindre» (Vidéo supplémentaire 1) ou «jouer avec la figure» (Tableau 1 ). Ce dernier comportement était spécifique à cette étape.
Figure 4.
Erreur moyenne et standard de fréquence pour le comportement «regarder la forme» réalisé par le lamantin avant (étape 2, en noir) et après (étape 4, en gris) entraînement. Test de Wilcoxon, *: P <0,05, NA: non disponible, NS: non significatif.
Lors de l'étape 2, aucune différence statistique (test de Friedman: df = 3, F = 6,81, p = 0,78) n'a été observée par rapport au temps passé devant chaque forme (moyenne en min ± SE; carré: 0,40 ± 0,30; triangle : 0,50 ± 0,20; disque: 0,60 ± 0,30; hexagone: 1,40 ± 0,70).
Nous avons conclu que Daniel était plus attiré par l'hexagone et le triangle mais moins attiré par le carré, que nous avons donc décidé d'utiliser pour l'entraînement. La présence de formes géométriques a suscité une augmentation notable des activités de Daniel.
3.2. Formation (étape 3)
Pendant la formation, Daniel a affiché les mêmes comportements que les étapes 1 et 2 mais aussi de nouveaux. Daniel a affiché un comportement «d'alimentation» (vidéo supplémentaire 2) lorsqu'il a mangé la laitue (tableau 1). Lorsque la laitue était sur la face ouverte de la forme, après avoir mangé, il «se reposait au fond de l'eau»; ce comportement ne s'est produit qu'au cours de cette étape (étape 3a). Daniel a également «mordu la silhouette» lors de l'étape avec de la laitue sur la face ouverte (étape 3a) et sur la face cachée (étape 3b). Un nouveau comportement «nager près de la silhouette» est apparu pendant l'entraînement et s'est produit pendant les deux phases. Le comportement «nager près de la figure» a été enregistré lorsque Daniel nageait normalement mais clairement à côté des formes (vidéo supplémentaire 3), ou spécifiquement à côté d'une seule forme. Lorsque la laitue était sur la face cachée de la figure, Daniel affichait de nouveaux comportements tels que «mordre le filet», «regarder derrière la forme» et «se reposer sur la figure». Au cours de l'étape 4, nous avons observé une augmentation impressionnante de l'activité.
Le temps qu'il a fallu à Daniel pour atteindre la place lorsque la laitue était suspendue devant elle était de 1,26 ± 0,70 min (moyenne ± SE). Lorsque la nourriture était suspendue derrière la forme, le temps enregistré était de 6,22 ± 4,00 min (moyenne ± SE). Malgré une nette tendance à mettre plus de temps à atteindre la forme lorsque la nourriture était en retard, aucune différence significative n'a été observée (test des paires appariées de Wilcoxon: Z = 1,82, p = 0,06). De plus, aucune différence significative n'a été observée (test des paires appariées de Wilcoxon: Z = 0,94, p = 0,34) dans le temps nécessaire pour commencer à manger la laitue devant la figure (moyenne ± SE: 5,00 ± 3,00 min) ou derrière (moyenne ± SE: 8,40 ± 1,20 min) (Vidéo supplémentaire 2). Le temps nécessaire pour consommer la laitue était identique (test des paires appariées de Wilcoxon:Z = 1,09, p = 0,27) quelle que soit la position de la laitue sur la forme (devant: 2,10 ± 0,70 min; derrière: 1,00 ± 0,30 min).
3.3. Après l'entraînement (étape 4)
Au cours de cette étape, Daniel a affiché les mêmes comportements que pendant l'entraînement, sauf pour «se nourrir» (Tableau 1). Le comportement «repos sur la forme», lorsque Daniel utilisait la figure comme support de sa tête et cessait de bouger, a été observé deux fois et n'était associé qu'au carré. Le comportement de «regarder derrière la figurine», lorsque Daniel a utilisé son nez pour pousser la figurine par derrière et l'inspecter, s'est produit avec le carré (5 observations) et une fois avec l'hexagone. Le comportement de «mordre la forme», lorsque Daniel mordit le bord de la forme, fut observé une fois sur l'hexagone. Le comportement «nager près de la forme» s'est produit pour toutes les formes (nombre d'observations: carré: 62, triangle: 39, disque: 43 et hexagone: 46) avec une nette préférence pour le carré, le chiffre le moins préféré avant l'entraînement (Vidéo supplémentaire 3). Au cours de cette étape, Daniel a également présenté une activité importante, comparable à celle présentée lors de la formation. Deux comportements semblent plus apparents lors de cette étape: «nager près de la forme» et «regarder la forme». Ces deux comportements ainsi que le comportement «regarder derrière la forme» montrent clairement que le lamantin recherche activement de la nourriture, en particulier sur le carré.
En regroupant les résultats pour quatre formes utilisées lors de l'expérience, la moyenne du nombre de chemins empruntés devant chaque forme avant l'entraînement (étape 2) (moyenne ± SE: 5,10 ± 0,60) est significativement plus faible (test des paires appariées de Wilcoxon: Z = 2,70, p = 0,006, N = 20) qu'après l'entraînement (étape 4) (moyenne ± SE: 9,50 ± 0,80). La figure 2 montre clairement une augmentation du nombre moyen de trajets pour chaque forme géométrique à l'exception du carré; il n'y a pas de différences statistiquement significatives entre avant et après la formation. Aucune différence n'a été observée pour les fréquences du nombre moyen de chemins entre les quatre formes après entraînement (test de Friedman: df = 3, F = 3.00, p= 0,40). Concernant ce comportement, il apparaît que Daniel a augmenté le nombre de chemins après l'entraînement, bien que les résultats ne permettent pas de faire une distinction entre les formes.
En regroupant les résultats pour toutes les formes utilisées pendant l'expérience, le nombre de comportements «regarder la forme» avant l'entraînement (moyenne ± SE: 0,40 ± 0,10 min) est significativement plus faible (test des paires appariées de Wilcoxon: Z = 3,40, p = 0,0007 , N = 20) qu'après (moyenne ± SE: 3,60 ± 0,70 min). La figure 4 montre clairement que le comportement «regarder la forme» n'est apparu qu'après l'entraînement avec le carré et jamais avant. En ce qui concerne les autres formes, la fréquence a augmenté de manière significative avec l'entraînement pour le disque et l'hexagone, mais pas pour le triangle. De plus, nous avons observé des différences significatives entre les fréquences des comportements «regarder la forme» (Figure 4) pour les quatre formes après entraînement (test de Friedman: df = 3, F = 12,40, p = 0,006) avec environ 7,5 comportements par répétition pour le carré alors que les fréquences pour les autres formes étaient toujours inférieures à 4.
En regroupant les résultats pour toutes les formes utilisées pendant l'expérience, la moyenne du comportement «visite» était significativement plus faible avant l'entraînement (moyenne ± SE: 1,10 ± 0,30) (test des paires appariées de Wilcoxon: Z = 3,00, p = 0,002, N = 20 ) qu'après (moyenne ± SE: 4,20 ± 0,70). La figure 3 montre que la fréquence observée pour le nombre de visites sur le carré a augmenté de manière significative après l'entraînement alors qu'aucune différence significative n'a été observée pour les trois formes restantes. Il y avait des différences significatives lors de la comparaison des fréquences de nombre de visites entre les quatre formes après l'entraînement (test de Friedman: df = 3, F = 10,40, p= 0,01) avec environ neuf visites par répétition pour le carré alors que le nombre de visites pour les autres formes était toujours inférieur à quatre (figure 3 ).
En regroupant les résultats pour toutes les formes utilisées pendant l'expérience, le temps passé sur les formes était significativement plus faible (test des paires appariées de Wilcoxon: Z = 2,02, p = 0,04) avant l'entraînement (moyenne ± SE: 2,50 ± 1,71 min) qu'après (moyenne ± SE: 10,30 ± 1,78 min). Le tableau 4 montre que cette différence est principalement due au fait que le lamantin est entraîné en utilisant de la nourriture sur le carré.
Tableau 4.
Temps moyen (min) passé par le lamantin dans des quadrants avec des formes lors des étapes 2 et 4 pour chaque forme
Forme Étape 2 (avant l'entraînement) Étape 4 (après l'entraînement) P
Carré 0.06 ± 0.04 7.20 ± 1.01 0.04
Triangle 1.60 ± 1.10 1.00 ± 0.50 NS
Disque 0.16 ± 0.05 0.80 ± 0.37 NS
Hexagone 0.78 ± 0.56 1.30 ± 0.30 NS
Test de Wilcoxon, NS = non significatif.
Tous ces résultats ont montré que l'entraînement modifiait le comportement de Daniel devant les formes, avec une tendance à une fréquence plus élevée ou à l'apparition de nouveaux comportements, en particulier concernant le carré.
3.4. Observations en dehors de la zone fermée (en lagune ouverte, étape 5)
Nous avons effectué deux observations en dehors de la zone fermée où les étapes expérimentales ont été effectuées (étape 1 à étape 4). Lors de la première observation, Daniel a effectué trois fois le comportement «regarder la forme» (Vidéo supplémentaire 4): deux fois pour le carré et une fois pour l'hexagone. Daniel a également exécuté le comportement «regarder derrière la forme» une fois sur le carré. Dans ce cas, il a touché la forme avec son nez pendant environ deux minutes. Après ces interactions, Daniel a quitté les zones de forme et n'est pas revenu pendant une heure d'observation. L'activité humaine dans le lagon l'a clairement dérangé une fois qu'il était dans la zone ouverte. Au cours de la deuxième observation, Daniel a été attiré par un petit bateau, puis a nagé vers le disque pour «regarder la forme», mais par la suite, il n'a passé plus de temps avec aucune des formes.
3.5. Observations après 3 et 12 mois (étape 6)
Lorsque les quatre formes sans nourriture ont été submergées dans la zone fermée, 3 et 12 mois après la formation, Daniel a seulement visité et interagi avec le carré (Vidéo supplémentaire 5). L'animal a nagé à plusieurs reprises près et autour de la place, regardant derrière l'objet pendant 3 et 12 mois après l'entraînement. Dans les deux observations, Daniel a également touché les deux faces du carré avec ses lèvres comme s'il en mangeait.
4. Discussion
Daniel était clairement attiré par les indices visuels utilisés dans notre expérience, avant même que l'une des formes géométriques ne soit associée à la nourriture. La présence de nouveaux objets a entraîné un changement évident dans le comportement du lamantin avec des modèles de comportement plus axés sur les objets et un niveau d'activité plus élevé, en particulier le mouvement. Ces résultats coïncident avec les réactions observées chez l'animal, par exemple chez la souris, lorsque les signaux visuels ne sont pas associés à la nourriture ou à l'eau [ 33 ]. L'attraction pour les objets était particulièrement élevée pour l'hexagone et le triangle, démontrant que Daniel avait une préférence pour ces formes. Il est difficile de savoir si cette préférence pour un hexagone et un triangle est due à des expériences antérieures avec des objets de forme géométrique similaire. Études sur les dauphins [ 9] a démontré que lorsque de nouveaux objets sont introduits dans la piscine, les dauphins sont visuellement attirés par eux, manipulent certains d'entre eux et peuvent avoir leur objet préféré. Un processus similaire pourrait avoir lieu dans cette étude avec de nouvelles formes, bien que le carré ne soit pas attrayant avant l'entraînement, peut-être parce que ce n'était pas une nouvelle forme pour Daniel. Cependant, nous n'avons aucune preuve de formes particulières avec lesquelles le lamantin était en contact avant ou après l'entraînement car Daniel vivait toujours dans les mêmes conditions. Daniel a visiblement perçu les formes dans l'eau, orientant son chemin vers elles, et les inspectant à travers des visites et un comportement de «regarder la forme». Cette capacité d'inspection à longue distance de gros objets a également été décrite par d'autres auteurs [ 27 , 34] bien que les lamantins semblent avoir moins de capacité visuelle que les autres mammifères aquatiques. Dans notre étude, il semble que les lamantins peuvent utiliser la vision pour explorer leur environnement sous l'eau. Ce résultat coïncide avec les observations précédentes [ 35 ] pour les distances longues et intermédiaires chez deux lamantins, qui présentaient une disparité des résultats entre les sujets car un individu ne présentait pas d'acuité visuelle à courte distance. Cependant, les lamantins ne vivent pas toujours en eau claire et il serait intéressant d'étudier si cette capacité est la même pour les individus en eau trouble et si les lamantins sont capables de répondre à des stimuli «hors de l'eau».
Après la formation, Daniel a clairement répondu à la forme associée à la nourriture. Le comportement «jouer avec la forme» n'a été observé qu'avant l'entraînement mais a disparu après que le lamantin ait associé la nourriture au carré. Nous suggérons qu'après l'association entre la nourriture et les formes, en particulier le carré, les formes géométriques ont acquis une signification particulière et Daniel a ajusté son comportement en fonction de la possibilité de présence d'aliments. Daniel a clairement montré sa capacité à associer rapidement des indices visuels à la possibilité de manger et à distinguer les formes géométriques. Cette affirmation est étayée par le fait qu'il a montré de nouveaux comportements après l'entraînement, associés à la recherche de la présence d'aliments dans les formes, avec un comportement de «regarder derrière la forme».31 , 34 ]. Nous ne nous attendions pas à ce qu'après trois mois, un lamantin soit toujours capable de discriminer et Daniel a toujours manifesté visiblement une attirance pour la place. Étonnamment, ce comportement s'est répété après 12 mois, accompagné d'un comportement typique de recherche alimentaire et de manipulation d'outils. Bien que ces résultats soient nouveaux pour les lamantins, la possibilité d'une mémoire à long terme est courante chez les mammifères. Par exemple, un border-collie nommé «Chaser» peut mémoriser le nom de plus d'un millier d'objets [ 36 ], et il y a des grands dauphins qui peuvent mémoriser les caractéristiques d'autres dauphins pendant des décennies [ 37 ]. Cette capacité de mémoire à long terme n'est pas surprenante si l'on considère que les lamantins phylogénétiquement sont des parents de l'éléphant [ 38 ,39 ] qui est connue pour son excellente mémoire [ 40]. En outre, la capacité d'un lamantin à utiliser les informations apprises en dehors de la zone fermée, même si les résultats sont limités, pourrait indiquer la capacité de généraliser les informations en tant que signaux qui sont utilisés pour trouver des ressources dans une zone naturelle. Dans ce contexte, lorsque les lamantins sont entraînés avant d'être relâchés, il peut être important de ne pas les entraîner avec des formes ou des objets qui pourraient les attirer vers des activités ou des constructions humaines, mais plutôt utiliser des éléments naturels trouvés dans la nature. Cependant, dans le cas de Daniel, élevé et imprimé par des humains nous considérons que ce n'est pas un réel problème car ce lamantin est toujours à la recherche de contacts humains. Cette capacité, qui consiste à utiliser des informations dans différents contextes, a déjà été décrite chez des oiseaux et plusieurs mammifères qui peuvent utiliser des informations apprises dans leur recherche de zones de nourriture ou de refuge [41 ]. Plusieurs travaux rapportent que les lamantins sont capables de parcourir de grandes distances à la recherche de nourriture, d'eau douce, d'eau chaude ou d'un partenaire sexuel [ 22 ]. La capacité de mémoriser et d'associer la nourriture à des signaux environnementaux a été observée chez plusieurs espèces d'invertébrés [ 12 , 15 ] et de vertébrés [ 41 ], et est décrite comme une stratégie adaptative pour optimiser la recherche de nourriture [ 12 , 42 ]. Compte tenu de nos résultats, il est raisonnable de suggérer que les lamantins peuvent apprendre un large éventail d'informations, leur permettant d'augmenter leur capacité de subsistance dès la naissance, en raison de fortes interactions mère-veau, d'interactions avec d'autres congénères [ 25] et les expériences individuelles ou sociales. On s'attend à ce que si un lamantin est capable d'utiliser des informations visuelles sous l'eau pendant la recherche de nourriture, les lamantins peuvent également utiliser des informations visuelles de leur environnement, sous ou hors de l'eau, lors de leurs déplacements pouvant couvrir de grandes distances [ 43 ]. Cependant, des études complémentaires sont nécessaires pour évaluer cette possibilité.
En raison de leur apparence et de leur lenteur à nager, les lamantins sont généralement considérés comme lents à comprendre ou peu intelligents par rapport aux autres mammifères aquatiques. Cette vision du lamantin devrait évoluer à la lumière de cette première étude formelle sur leur capacité d'apprentissage et de mémoire. En fait, leur capacité cognitive, avec leurs spécificités, peut être plus proche de celle d'autres mammifères aquatiques dotés d'un grand cerveau [ 30]. Certaines applications pratiques potentielles de nos résultats méritent d'être mentionnées. Premièrement, après cette étude pionnière, d'autres études sur les capacités d'apprentissage des lamantins sont essentielles, en utilisant différents indices pour tester différents aspects de leur environnement, tels que les exigences sociales. Deuxièmement, la possibilité d'utiliser des repères visuels pour entraîner les lamantins pourrait être utilisée pour les gérer, les entraîner et les préparer sans interaction humaine pour la libération dans la nature et les programmes de réintroduction. Les possibilités d'enrichissement sensoriel et environnemental pour le bien-être du lamantin dans un environnement d'aquarium pourraient être associées à la possibilité de préparer certains animaux à la réintroduction dans un nouvel environnement. En réalité,26 ]. En outre, l'association entre les objets et la nourriture qui a suscité plus d'activités, la manipulation d'objets et la recherche active sur les aliments utilisant la mémoire à court et à long terme ou des indices dans différents contextes peut être un moyen pratique d'améliorer le bien-être des lamantins, compte tenu de l'importance de l'utilisation de la cet objectif [ 4 , 6 ]. Nous espérons que des études similaires contribueront à améliorer les programmes de gestion des lamantins, ainsi qu’à encourager de nouvelles recherches sur le comportement des lamantins dans le but d’améliorer leur bien-être.
Les animaux relâchés dans la nature se sont souvent révélés incapables de s'adapter aux nouvelles conditions, car les animaux élevés dans des conditions non naturelles doivent se renseigner sur les prédateurs, comment trouver de la nourriture et comment survivre dans différentes conditions naturelles [ 44 ]. L'importance vitale de l'apprentissage et de la préparation avant d'être relâché est bien documentée chez les invertébrés [ 13 , 15 ] et les vertébrés [ 45 ]. Dans le cas des lamantins, plusieurs tentatives de réintroduction n'ont pas été aussi fructueuses que prévu [ 46]; par conséquent, nous considérons que cette étude pourrait contribuer à améliorer la préparation avant la remise en liberté de ces mammifères aquatiques en voie de disparition et ainsi augmenter les chances de réussite du lâcher et de survivre dans leur habitat naturel. Par exemple, les lamantins doivent trouver de l'eau douce et des produits alimentaires spécifiques dans de vastes zones; le fait que les veaux vivent avec leur mère pendant de longues périodes où ils peuvent apprendre l'emplacement des sources de nourriture et d'eau, peut être clairement lié à des indices visuels et autres associés à la présence de ressources. Cette nécessité d'apprentissage doit être prise en compte dans les programmes de réintroduction.
Cette étude a fourni de nouvelles informations sur les capacités d'apprentissage et la mémoire à long terme chez les lamantins. Cependant, la recherche s'est concentrée sur un lamantin solitaire élevé par l'homme et il serait intéressant de développer des études similaires avec des lamantins sauvages, en utilisant des éléments plus naturels. Dans ce contexte, notre étude montre également comment les lamantins augmentent leur activité et le nombre de comportements en présence de nouveaux objets. Par conséquent, l'introduction de nouveaux objets pour stimuler l'apprentissage associatif avec des repères visuels fournit une nouvelle méthode intéressante pour augmenter le bien-être des lamantins en captivité. De plus, nos découvertes peuvent contribuer à la préparation et à la réintroduction réussie du lamantin dans son environnement naturel. Cette étude pionnière offre de nouvelles perspectives d'études comportementales sur les lamantins, en particulier celles associées à la cognition, à la gestion et au bien-être.
Les conflits d'intérêts
Les auteurs déclarent ne pas avoir de conflits d'intérêts concernant cet article.
Remerciements
Nous remercions le Ministère du Développement Urbain et de l'Environnement pour les installations prévues pour la réalisation des travaux au Centre de soins et de réadaptation des mammifères aquatiques. Un grand merci à CONACYT et ECOSUR pour avoir fourni une bourse d'études MSc à Lara-Sánchez. Nous remercions Olivier Velázquez pour le travail vidéo ainsi que M. Eladio Juarez et sa famille pour le soutien et les services offerts.
Données supplémentaires
Des informations complémentaires (vidéos) pour cet article sont disponibles sur le site Web de la revue sous https://doi.org/10.5802/crbiol.6 ou auprès de l'auteur.
Bibliographie
F. Mery Natural variation in learning and memory, Curr. Opin. Neurobiol., Volume 23 (2013) no. 1, pp. 52-56
J. M. Pearce; M. E. Bouton Theories of associative learning in animals, Annu. Rev. Psychol., Volume 52 (2001), pp. 111-139
A. Dickinson Associative learning and animal cognition, Phil. Trans. R. Soc. B, Volume 367 (2012), pp. 2733-2742
F. E. Clark Marine mammal cognition and captive care: a proposal for cognitive enrichment in zoos and aquariums, J. Zoo Aqua. Res., Volume 1 (2013) no. 1, pp. 1-6
H. Whitehead; L. Rendell; R. W. Osborne; B. Würsig Culture and conservation of non-humans with reference to whales and dolphins: review and new directions, Biol. Conserv., Volume 120 (2004), pp. 427-437
F. E. Clark; L. J. Smith Effect of a cognitive challenge device containing food and non-food rewards on chimpanzee well-being, Am. J. Primatol., Volume 9999 (2013), pp. 1-10
C. L. Meehan; J. A. Mench The challenge of challenge: can problem solving opportunities enhance animal welfare?, Appl. Anim. Behav. Sci., Volume 102 (2007), pp. 246-261
R. C. Newberry Environmental enrichment: increasing the biological relevance of captive environments, Appl. Anim. Behav. Sci., Volume 44 (1995), pp. 229-243
F. Delfour; H. Beyer Assessing the effectiveness of environmental enrichment in bottlenose dolphins (Tursiops truncatus), Zoo Biol., Volume 31 (2012), pp. 137-150
M. S. Szokalskia; C. A. Litchfielda; W. K. Fosterb Enrichment for captive tigers (Panthera tigris): current knowledge and future directions, Appl. Anim. Behav. Sci., Volume 139 (2012), pp. 1-9
D. L. Wells Sensory stimulation as environmental enrichment for captive animals: a review, Appl. Anim. Behav. Sci., Volume 118 (2009), pp. 1-11
Y. Hénaut; S. Machkour-M’Rabet; J. P. Lachaud The role of learning in risk-avoidance strategies during spider–ant Interactions, Anim. Cogn. (2014), pp. 185-195
Y. Hénaut; C. Alauzet; A. Ferran; T. Williams Effect of nymphal diet on adult predation behavior in Orius majusculus (heteroptera: Anthocoridae), J. Econ. Entomol., Volume 93 (2000), pp. 252-255
S. Wanatabe Strategies of spatial learning for food storing in scrub jays, J. Ethol., Volume 23 (2005), pp. 181-187
Y. Hénaut; C. Alauzet; D. Dargagnon; M. Lambin Visual learning larval Orius majusculus a polyphagous predator, Entomol. Exp. Appl., Volume 90 (1999), pp. 103-107
C. W. Werner; G. Rehkämper Discrimination of multidimensional geometrical shapesshapes by chickens: categorization and pattern-learning, Anim. Cogn., Volume 2 (1999), pp. 27-40
L. Martinez; T. Matsuzawa Visual and auditory conditional position discrimination in chimpanzees (Pan troglodytes), Behav. Processes, Volume 82 (2009), pp. 90-94
J. Langbein; K. Siebert; G. Nuernberg Concurrent recall of serially learned visual discrimination problems in dwarf goats (Capra hircus), Behav. Processes, Volume 79 (2008), pp. 156-164
H. E. Harley Whistle discrimination and categorization by the Atlantic bottlenose dolphin (Tursiops truncatus): a review of the signature whistle framework and a perceptual test, Behav. Processes, Volume 77 (2008), pp. 243-268
A. Kilian; S. Y. L. Von Fresen; O. Güntürkün A bottlenose dolphin discriminates visual stimuli differing in numerosity, Learn. Behav., Volume 31 (2003), pp. 133-142
J. Z. Abramson; V. Hernández-Lloreda; J. Call; F. Colmenaresa Relative quantity judgments in the beluga whale (Delphinapterus leucas) and the bottlenose dolphin (Tursiops truncatus), Behav. Processes, Volume 96 (2013), pp. 11-19
D. S. Hartman Ecology and Behavior of the Manatee (Thrichechus manatus) in Florida, Special Publication no. 5, American Society of Mammalogist, 1979, pp. 1-153
J. K. Koelsch (“The seasonal occurrence and ecology of Florida manatees (Trichechus manatus latirostris) in coastal waters near Sarasota”, Florida, MS dissertation, University of South Florida, 1997)
J. Y. Harper; B. A. Schulte Social interactions in captive female Florida manatees, Zoo Biol., Volume 24 (2005), pp. 135-144
Y. Hénaut; S. Becerra-López; S. Machkour-M’Rabet; B. Morales-Vela; P. Winterton; F. Delfour Activities and social interactions in captive antillean manatees in Mexico, Mammalia, Volume 74 (2010), pp. 141-146
D. G. Anzolin; P. S. M. De Carvalho; P. C. Vianajr; I. C. Normande; A. Da Silva-Souto Stereotypical behaviour in captive West Indian manatee (Trichechus manatus), J. Mar. Biol. U.K., Volume 94 (2014), pp. 1133-1137
R. L. Reep; R. K. Bonde The Florida Manatee: Biology and Conservation, University Press of Florida, Gainesville, 2006, pp. 1-189
J. E. Reynolds III; D. K. Odell Manatees and Dugongs, Facts on File, Inc., New York, 1991 (192 p.)
C. D. Marshall; G. D. Hurt; U. M. Edmond; D. L. Hallin; R. L. Reep Prehensile use of penoral bristles during feeding and associated behaviors of the Florida Manatee (Trichechus manatus latirostris), Mar. Mam. Sci., Volume 14 (1998), pp. 274-289
D. K. Sarko; J. I. Johnson; R. C. Switzer III; W. I. Welker; R. L. Reep Somatosensory nuclei of the manatee brainstem and thalamus, Anat. Rec., Volume 290 (2007), pp. 1138-1165
D. Dos Santo-Lima; J. E. Vergara-Parente; R. J. Young; E. Paszkiewicz Training of antillean manatee Tricherus manatus manatus Linnaeus 1758, as a management technique for individual welfare, Lat. Am. J. Aqua. Mam., Volume 4 (2005), pp. 61-68
G. Bennett; J. R. Galef; K. N. Laland Social learning in animals: empirical studies and theoretical models, Bioscience, Volume 55 (2005), pp. 489-499
C. J. Heyser; A. Chemero Novel object exploration in mice: not all objects are created equal, Behav. Processes, Volume 89 (2012), pp. 232-238
E. R. Gerstein The manatee mind: discrimination training for sensory perception testing of West Indian Manatees (Tricherus manatus), Mar. Mam. Public Display Res., Volume 1 (1994), pp. 10-21
G. B. Bauer; D. E. Colbert; J. C. Gaspard III; B. Littlefiled; W. Fellner Underwater visual acuity of Florida manatees (Trichechus manatus latirostris), Int. J. Comput. Psychiatry, Volume 16 (2003), pp. 130-142
J. W. Pilley; A. K. Reid Border collie comprehends object names as verbal referents, Behav. Processes, Volume 86 (2011), pp. 184-195
J. N. Bruck Decades-long social memory in bottlenose dolphins, Proc. R. Soc. Lond. B, Volume 280 (2013) (20131726)
W. E. Rainey; J. M. Lowenstein; V. M. Sarich; D. M. Magor Sirenian molecular systematics - including the extinct Steller’s sea cow (Hydrodamalis gigas), Naturwissenschaften, Volume 71 (1984), pp. 586-588
P. D. Waters; G. Dobigny; P. J. Waddell; T. J. Robinson Evolutionary history of LINE-1 in the major clades of placental mammals, PLoS ONE, Volume 2 (2007) (e158)
B. L. Hart; L. A. Hart; N. Pinter-Wollman Large brains and cognition: where do elephants fit in?, Neurosci. Biobehav. Rev., Volume 32 (2008), pp. 86-98
S. Healy; S. de Kort; N. S. Clayton The hippocampus spatial memory and food hoarding: puzzle revisited, Trends Ecol. Evol., Volume 1 (2005), pp. 17-22
S. Healy; C. M. Jones Animal learning and memory: an integration of cognition and ecology, Zoology, Volume 4 (2002), pp. 321-327
D. N. Castelblanco-Martínez; J. Padilla-Saldívar; H. A. Hernández-Arana; D. H. Slone; J. P. Reid; B. Morales-Vela Movement patterns of Antillean manatees in Chetumal Bay (Mexico) and coastal Belize: a challenge for regional conservation, Mar. Mam. Sci., Volume 29 (2013), pp. 166-182
W. T. Greenough; J. M. Juraska Experience induced changes in brain fine structure: their behavioral implications, Development and Evolution of Brain Size: Behavioral Implications (M. E. Hahen; C. Jensen; B. C. Dudek, eds.), Academic Press, New York, 1979, pp. 263-294
A. Vargas; D. Biggins; B. Miller Etología aplicada al manejo de especies amenazadas: el caso del turón de patas negras (Mustela nigripes), Etología, Volume 7 (1999), pp. 33-39
A. S. Griffin; C. S. Evans; D. T. Blumstein Learning specificity in acquired predator recognition, Anim. Behav., Volume 14 (2001), pp. 1317-1326